Lichen
Modèle:Confusion Modèle:Voir homonymes
{kind=link}
.jpg){kind=link}
Le lichen (Modèle:MSAPI), appelé aussi champignon lichénisé ou champignon lichénisant, est un organisme composite qui résulte d'une symbiose permanente entre au moins un champignon hétérotrophe appelé mycobionte, et des cellules microscopiques photoautotrophes, possédant de la chlorophylle, nommées photobiontes. Le lichen forme un holobionte constitué d'une multitude de symbiontes avec des stratégies évolutives différentes, parfois antagonistes, parfois complémentaires. Le mycobionte est prépondérant dans la plupart des genres. Le photobionte s'appelle phycobionte lorsque le partenaire est une algue verte (ce qui donne la dénomination de chlorolichen ou phycolichen), cyanobionte ou bactériobionte lorsqu'il est une cyanobactérie (ce qui donne la dénomination de cyanolichen). Les lichens sont classés dans le phylum des Fungi.
Les lichens génèrent un appareil végétatif composé à 90 % par le champignon, le thalle, qui se développe lentement à la surface de supports variés, y compris dans des milieux souvent hostiles (exposition à la sécheresse, à de fortes températuresModèle:Etc.).
Depuis 2011, de nombreuses découvertes mettent en évidence que la symbiose lichénique implique de multiples partenaires (levures, bactéries, protistes, virus).
La symbiose multipartenariale résulte d'une association, appelée lichénification ou lichénisation. L'inverse, c'est-à-dire une algue macroscopique hébergeant un champignon microscopique, est une mycophycobiose.
L'étude des lichens est appelée « lichénologie ».
Les lichens sont la forme de vie dominante sur 6 à 8 % de la surface terrestre<ref>Modèle:Article.</ref>,<ref>Modèle:Article.</ref>, cette composante importante de l'écosystème terrestre étant principalement dû aux lichens terricoles du genre Cladonia (lichens des rennes) qui peuvent couvrir plus de 97 % des surfaces à l'intérieur des forêts boréales ouvertes et y former des tapis de 10 à Modèle:Unité d'épaisseur<ref>Modèle:Article.</ref>. Les lichénologues ont inventorié Modèle:Unité<ref>Modèle:Lien web.</ref>. Près de 100 nouvelles espèces sont découvertes chaque année et il en resterait 8 000 à découvrir<ref>Modèle:Article.</ref>. Les plus vieux fossiles de lichens datent du Cambrien<ref>Modèle:Article.</ref>.
Étymologie
Le mot « lichen » vient du grec Modèle:Grec ancien, « dartre, cal, plante parasite ». Il est dérivé de Modèle:Grec ancien, « lécher », qui remonte à la même racine indo-européenne que lécher : la maladie, comme l'organisme semblant lécher son support. Historiquement, le premier sens de lichen est celui d'« éruption sur la peau », conservé par la dermatologie moderne dans une acception moins générique. Le sens courant en botanique, attesté depuis 1546, se désigne les structures épiphytes dont les lichens encroûtants, présentés comme des dartres<ref>Modèle:Ouvrage.</ref>.
Historique
Le terme de lichen est utilisé la première fois, par Théophraste, au Modèle:Lien siècle av JCModèle:Vérification siècle Dans le Livre III de son Histoire des plantes, le philosophe grec dit que les lichens naissent de l'écorce, et non d'un œil, et les compare à des guenilles<ref>Théophraste, Histoire des plantes, Livre III, chap. 11 Des signes du temps.</ref>. Il désigne alors des hépatiques et leur attribue des vertus médicinales, les lichens réellement décrits par Théophraste étant nommés autrement<ref>Modèle:Ouvrage</ref>.
Jusqu'au milieu du Modèle:Lien siècleModèle:Vérification siècle, les naturalistes les classent dans la catégorie des mousses, des champignons ou, selon Linné et ses disciples, dans la catégorie des algues au sein des cryptogames. Ces naturalistes considèrent les lichens du sol souvent comme des Modèle:Citation<ref>Modèle:Harvsp.</ref>. Il faut attendre l'avènement de microscopes performants pour que les interactions biotiques dans ces organismes soient mises en évidence. En 1867, le botaniste suisse Simon Schwendener est le premier à considérer qu'il a une double nature, sous forme de parasitisme<ref>Modèle:Article.</ref>. Beaucoup de lichénologistes reconnus rejettent d'abord cette « hypothèse duale des lichens », car le consensus est alors partagé sur le fait que tous les organismes vivants sont autonomes<ref>Modèle:Article.</ref>. Mais d'autres éminents biologistes, comme Anton de Bary ou Albert Bernhard Frank, sont moins prompts à repousser les idées de Schwendener, et ce concept s'étend bientôt à d'autres domaines d'étude, comme les agents pathogènes. Albert Bernhard Frank propose le terme de symbiotismus en 1877<ref>Modèle:Article.</ref>, terme peu à peu accepté par la communauté scientifique à la suite des travaux d'Anton de Bary qui donne la définition la plus large de la symbiose<ref>Modèle:Article.</ref>. L'« hypothèse duale » est démontrée expérimentalement par le professeur allemand Rees en 1871, l'algologue Bornet en 1873 (il réussit à montrer que les haustoriums fongiques pénètrent dans les cellules d'algues) et le botaniste Bonnier en 1886 et 1889<ref>Modèle:Ouvrage.</ref>.
Au Modèle:S mini- siècleModèle:Vérification siècle, la systématique moderne classe les lichens dans le phylum des Fungi car seul le mycobionte assure la reproduction sexuée. Depuis 2011, les techniques modernes de biologie moléculaire du type métagénomique et pyroséquençage montrent que la symbiose lichénique implique de multiples partenaires (levures, bactéries, protistes, virus)<ref name="MDPI">Modèle:Article.</ref>.
Symbioses
{kind=link}
{kind=link}
.png){kind=link}
_(8558627918).jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Généralités sur les partenaires
Les lichens font partie de la biodiversité négligée ; bien qu'ils soient peu recherchés lors des inventaires fongiques, une centaine de nouvelles espèces sont décrites chaque année. En 2007, 18 882 espèces de lichens sont décrites<ref>Modèle:Article.</ref>. En Europe occidentale, le nombre d'espèces est compris entre 50 et 75 % du nombre de phanérogames. En France, on compte environ Modèle:Unité pour Modèle:Unité<ref>Modèle:Harvsp.</ref>.
La spécificité d'association entre photobionte et mycobionte(s) peut être étroite ou large mais la plupart des lichens sont modérément spécifiques : un même mycobionte peut s'associer à différentes espèces de phytobionte. Les différentes associations sont<ref>Modèle:Chapitre.</ref> :
- dans 85 % des cas, une (ou des) algue(s) associée(s) à un ou plusieurs champignons ;
- dans 10 % des cas, une cyanobactérie associée à un champignon ;
- dans 5 % des cas, les 3 types de partenaires sont associés : dans cette symbiose tripartite, la cyanobactérie fixatrice d'azote est généralement confinée dans des compartiments fongiques appelés Modèle:Lien<ref group=alpha>{{#invoke:Langue|indicationDeLangue}} Structure anatomique schématique de lichens montrant des céphalodies. Les cyanobactéries y réalisent la fixation de l'azote dans des cellules spécialisées, généralement un peu plus grandes que les cellules végétatives, les hétérocystes, grâce à une enzyme particulière, la nitrogénase. Cf Modèle:Article.</ref>, structures internes ou externes<ref>Modèle:Article.</ref>.
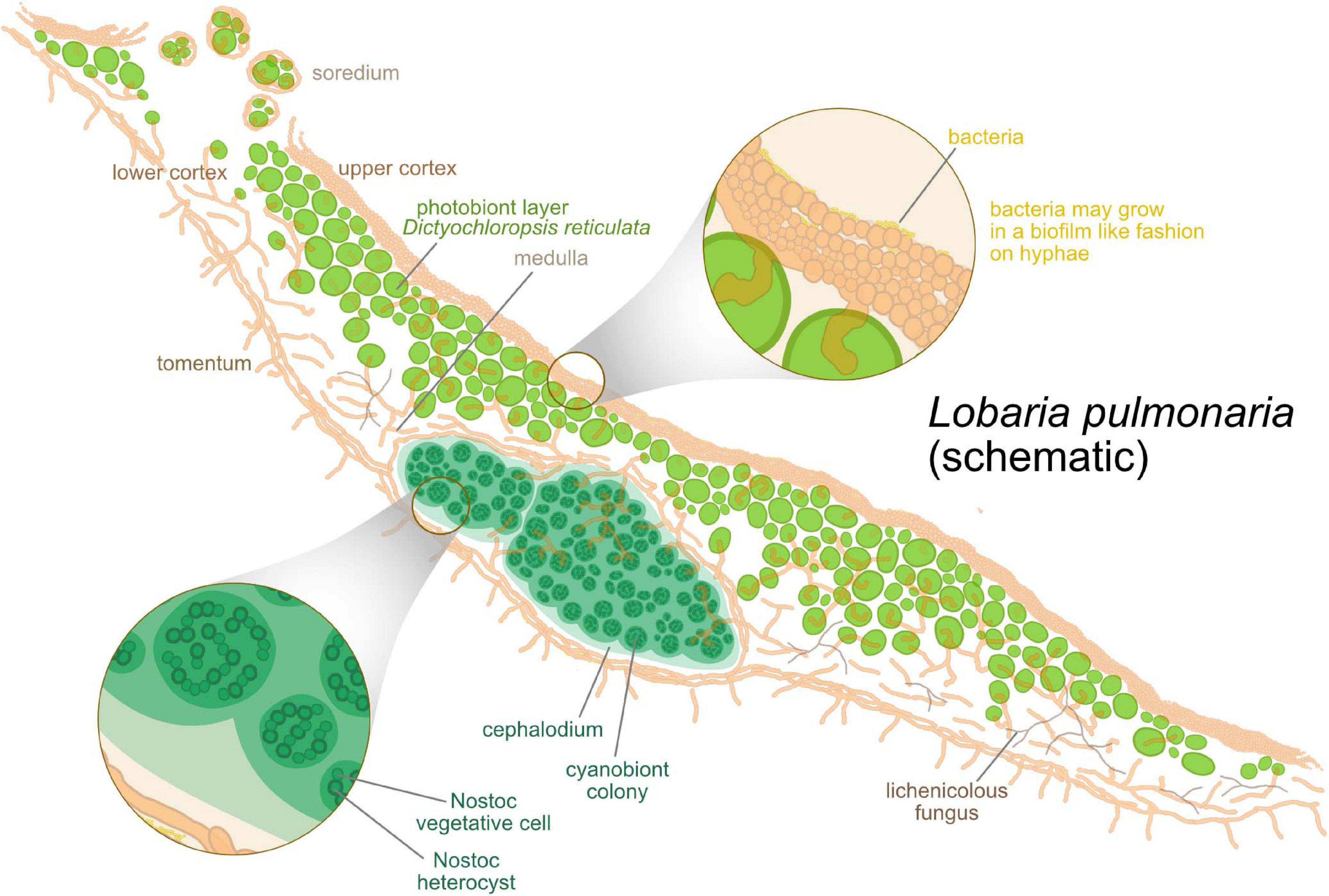{kind=link}
Mycobiontes
Les mycobiontes impliqués dans la symbiose lichénique (ascomycète, basidiomycète ou deutéromycète) représentent environ un cinquième de l'ensemble des champignons actuellement connus. On rencontre le plus souvent des « ascolichens » dans lesquels l'ascomycète est toujours un symbiote obligatoire : 40 % des espèces d'ascomycètes ont adopté ce type de symbioseModèle:Sfn, dont principalement les Pezizomycotina<ref>Modèle:Article.</ref>.
Après plus de 140 ans durant lesquels on a cru que l'association était binaire (1 champignon + 1 algue), des chercheurs ont montré en 2016 qu'il faut en réalité dans la plupart des lichens un troisième partenaire (qu'on avait d'abord cru être un parasite des lichens) pour que l'association soit pérenne ; il s'agit d'une levure basidiomycète ; c'est elle qui est responsable de la forme du thalle du lichen et en partie de sa forme générale<ref name="2016spribille">Modèle:Article.</ref>. La découverte de ce troisième partenaire (la levure basidiomycète) est entre autres due à Toby Spribille assistant professeur à la faculté des sciences de l'Alberta et à l'équipe de McCutcheon<ref name="cifar">Modèle:Lien web.</ref>. Ces travaux furent initiés par la comparaison entre deux espèces de lichens , B. tortuosa et B. fremontii séparées par la production d'acide vulpinique. Après plusieurs tests effectués, l'hypothèse d'une différence d'expression de gènes à l'origine de l'acide vulpinique fut écartée. Ils ont fini par découvrir que cette production d'acide vulpinique jusqu'alors inexpliquée était en réalité due à la présence de levure dans le thalle du lichen<ref name="2016spribille"/>,<ref name="cifar" />. Cette association est durable, reproductible (elle donne naissance à de nouveaux individus, à la formation d'une nouvelle unité fonctionnelle) avec des bénéfices réciproques pour les partenaires, et entraîne des modifications morphologiques et physiologiques (ces dernières liées à des interactions génétiques entre les trois partenaires)<ref name="2016spribille" />. Les levures sont intégrées dans le cortex lichénique. Il y a une corrélation entre leur abondance et des variations autrefois inexpliquées du phénotype<ref name="2016spribille" />. Certaines lignées de basidiomycètes vivent en lien étroit avec certains lichens sur de vastes zones géographiques, souvent retrouvés sur les six continents<ref name="2016spribille" />. Le cortex lichénique est structurellement plus important qu'on ne le pensait ; il ne s'agit pas d'une simple zone de cellules différenciées d'ascomycètes, mais semble-t-il du lieu d'une symbiose (entre deux types différents de champignons) dans la symbiose (champignon-plante)<ref name="2016spribille" />.
Photobiontes
85 % des photobiontes sont des algues vertes (phycobionte de type chlorophytes) appartenant pour 98 % d'entre eux à la classe des Chlorophyceae qui regroupe la famille des Chlorococcaceae représentée pour plus de 60 % par l'ordre des Trebouxiales avec le genre Trebouxia (20 %), lichen souvent vert, et par l'ordre des Trentepohliales avec le genre Trentepohlia (lichen souvent orange-rouge, en raison de ces algues dont les gouttelettes lipidiques renferment des pigments caroténoïdes)<ref>Modèle:Ouvrage.</ref>. Il s'agit souvent de symbiotes facultatifs. Sur deux cents espèces au total, une seule espèce d'algue non verte existe (une xanthophycée chez des lichens aquatiques du genre Verrucaria).
Les champignons sélectionnent deux taxons de photobiontes algaux selon leur substrat et leur préférence écologique (exposition à la pluie et au soleil)<ref>Modèle:Chapitre</ref>.
10 % des espèces lichéniques possèdent un photobionte appartenant à la classe des Cyanobactéries. Ces symbiontes facultatifs sont des bactéries de forme unicellulaire ou filamenteuse, avec contenu cellulaire vert bleuâtre ou vert brunâtre, représentées par quatre familles : Chroococcales (Modèle:Lien), Pleurocapsales, Stigonematales et surtout les Nostocales (Modèle:Lien, Nostoc présent dans 3 % des lichens). 5 % des espèces lichéniques possèdent simultanément les deux types de photobiontes. Les partenaires autotrophes étant loin d'être tous identifiés, les chiffres présentés ici sont provisoires<ref>Modèle:Ouvrage.</ref>.
À noter que les microalgues des lichens ne sont pas les seuls organismes photosynthétiques à entrer dans un schéma de symbiose, des macroalgues le peuvent également (mycophycobioses).
Bactériobiontes
Les bactériobiontes sont des communautés bactériennes associées aux deux partenaires symbiotiques (photobionte et mycobionte). Ces espèces épiphytiques (biofilms sur le thalle) et endophytiques synthétisent des métabolites spécialisés bioactifs qui ont un rôle de protection vis-à-vis d'autres organismes pathogènes ou parasitiques<ref>Modèle:Article.</ref>.
Autres partenaires
De nombreuses découvertes mettent en évidence que la symbiose lichénique implique d'autres partenaires (protistes, virus…) qui s'intègrent dans un spectre d'interactions biotiques, allant du commensalisme et du mutualisme (organismes jouant un rôle dans la structuration du thalle et la modulation de la réponse du lichen à des facteurs environnementaux<ref name="MDPI" />), au parasitisme et à la pathogénicité<ref>Schéma représentant différentes interactions biotiques au sein des lichens. Cf Modèle:Article.</ref>.
{kind=link}
Anatomie et structure des thalles
{kind=link}
Deux types d'anatomie du thalle caractérisent le lichen<ref>Modèle:Harvsp.</ref> :
- lichen homomère ou homéomère dont les cellules du photobionte sont réparties uniformément dans le thalle (cas des genres Leptogium et Collema) ;
- lichen hétéromère dont les cellules du photobionte sont regroupées dans une couche assimilatrice.
La structure d'un lichen hétéromère est typiquement formée :
- d'un cortex supérieur constitué d'un synenchyme (tissu compact formé par les hyphes resserrés du champignon) ;
- d'une couche assimilatrice constitué des cellules du photobionte réparties dans le prosenchyme (tissu fibreux formé par les hyphes biens séparés du champignon) ;
- d'une médulle ou couche médullaire, prosenchyme très lâche sans photobionte ;
- d'un cortex inférieur lui aussi constitué d'un synenchyme ;
- de Modèle:Lien constituées de faisceaux d'hyphes plus ou moins soudées et recouvertes d'une gaine gélatineuse facilitant l'adhésion au substrat (les rhizines n'ont qu'un rôle d'ancrage, à la différence des racines chez les plantes vasculaires qui assurent leur nutrition hydrominérale).
Cette structure peut considérablement varier chez une même espèce de lichen selon son substrat, l'humidité.
Les lichénologues distinguent six types de lichens selon l'aspect global de leur thalle<ref>Modèle:Harvsp.</ref> :
- « lichen crustacé ou incrustant » (90 % des lichensModèle:Sfn) présentant un thalle hétéromère fortement plaqué<ref group=alpha>On ne peut le détacher de son substrat qu'en l'abîmant fortement.</ref> au support, formant une croûte. En fonction de sa position vis-à-vis du substrat, ce lichen peut être endosubstratique ou épisubstratique (épi- ou endolithique dans le cas d’une roche, épi- ou endophléode dans le cas d’un lichen corticole, épi- ou endogé pour un lichen terricole). Selon la forme du thalle, les lichénologues distinguent deux sous-types<ref>Modèle:Article.</ref> :
- lichen crustacé lobé au pourtour : thalle placodiomorphe (Modèle:Ex Fulgensia fulgens, Solenopsora candicans) ;
- lichen crustacé non lobé au pourtour : thalle continu, lisse, rugueux ou plissé, fendillé-aréolé, granuleux ou verruqueux, pulvérulent ou lépreux<ref group=alpha>Les thalles lépreux (exemples : Lepraria, Modèle:Lien) considérés comme primitifs, parviennent à constituer de grandes surfaces farineuses, surtout sur des substrats ombragés et protégés des eaux de ruissellement pour éviter leur lessivage.</ref> (Modèle:Ex Rhizocarpon geographicum, Acrocordia conoidea, Aspicilia farinosa, Rinodina confragosa, Lecidella carpathica) ;
- « lichen foliacé » présentant un thalle hétéromère non soudé sur toute sa surface (se détachant facilement du substrat), formant des lames souvent lobées comme de petites feuilles qui s'écartent un peu du support, présence d'haptères (crampons) ou de rhizines (fausses radicelles) sur leur face inférieure pour adhérer au substrat :
- lichen foliacé ombiliqué : dépression dénommée ombilic sur la face supérieure (Modèle:Ex Umbilicaria grisea, Umbilicaria polyphylla) ;
- lichen foliacé non ombiliqué : pas d'ombilic, lobes à disposition radiée (Modèle:Ex les Xanthoria, les Parmeliaceae comme Punctelia borreri, Parmelia omphalodes).
- « lichen fruticuleux » (du latin frutex, « arbrisseau ou buisson ») présentant un thalle adhérent au substrat par une surface réduite et formant des prolongements redressés, pendants ou étalés. Ces prolongements plus ou moins longs présentent trois formes :
- tiges rondes plus ou moins ramifiées (Modèle:Ex Alectoria ochroleuca, Usnea florida, Thamnolia vermicularis) ;
- lanières plates parcourues par des cannelures (Modèle:Ex Ramalina fraxinea) ;
- « lichen squamuleux » : squamules (petits compartiments ressemblant à des écailles) à la surface supérieure (Modèle:Ex genres Lecanora, Toninia, Rhizoplaca) ;
- « lichen complexe » ou « lichen composite » présentant un thalle primaire plus ou moins foliacé étalé sur le substrat, et sur lequel se développe un thalle secondaire fruticuleux, formé d'éléments (ramifiés, en forme de corne, d'entonnoir ou de trompette, type Modèle:Lien) se développant perpendiculairement au substrat (genre Cladonia et Stereocaulon vulcani) ;
- « lichen gélatineux » présentant un thalle à cyanobactérie (thalle foliacé, squamuleux, crustacé ou fruticuleux), noir et cassant à l'état sec, devenant gélatineux sous l'action de l'eau (Modèle:Ex genres Thyrea et Leptogium, Collema subnigrescens).
Les lichénologues distinguent parfois les « macrolichens » (forme des lichens squamuleux, foliacés ou fruticuleux), des « microlichens » (lichens crustacés granuleux, verruqueux, pulvérulents ou lépreux)<ref>Modèle:Ouvrage.</ref> ; les préfixes « macro » et « micro » ne se réfèrent donc pas ici à la taille du lichen, mais à la hauteur de son thalle et à la forme de croissance du lichen<ref>What is a lichen?, Australian National Botanical Garden.</ref>.
En ce qui concerne les relations entre le mycobionte et le phytobionte, on distingue trois cas de figure :
- l'hyphe du champignon se propage entre les cellules du nostoc (algue bleue, genre cyanobactérie) et dans son mucilage ;
- la formation d'un appressorium où les deux partenaires ont leurs parois en apposition, accolées l'une à l'autre avec une légère modification. Le contact est plus étroit et plus sophistiqué ;
- la formation d'un haustorium où le phytobionte finit par se trouver inclus dans la paroi du mycobionte. Ici, les modifications cytologiques sont grandes.
-
Microlichen « crustacé » du genre Rhizocarpon sur un rocher alpin, avec son hypothalle bien visible<ref group=alpha>À la périphérie du thalle et entre les aréoles, on peut observer l'hypothalle noir, appareil végétatif qui remplace le cortex inférieur dans de nombreux thalles crustacés. Cet hypothalle, formé d'hyphes parallèles à la surface du substrat dans lequel il émet des hyphes fixatrices, constitue le mycélium du champignon seul, sans les photobiontes. Dans les thalles crustacés très minces, plus spécialement dans les thalles endolithiques et endophléodes, l'hypothalle et le cortex inférieur font défaut, le cortex supérieur (réduit à quelques hyphes lâches) et la médulle s'amincissent, la structure tendant à devenir homomère. Cf Modèle:Ouvrage.</ref>.
-
Modèle:Lien, endolithe dont seules sont visibles les apothécies noires suivant des lignes de rupture de la roche.
-
Protoparmeliopsis muralis, lichen crustacé saxicole couvrant de nombreux substrats artificiels<ref group=alpha>Murs, béton, mortier, tuiles…</ref>.
-
Ramalina fastigiata, lichen fruticuleux, corticole sur arbres à feuilles caduques.
-
Lichen foliacé de Physcia.
{kind=link}
.jpg){kind=link}
.jpg){kind=link}
.jpg){kind=link}
.JPG){kind=link}
Reproduction et nutrition
.png){kind=link}
Reproduction
Le lichen a plusieurs modes de reproduction : reproduction végétative ou reproduction asexuée, et reproduction sexuée.
La reproduction végétative se fait par simple fragmentation du thalle (phénomène du bouturage) ou à l'aide d'organes spécialisés<ref>Modèle:Harvsp.</ref> qui s'en détachent :
- des isidies, protubérances de formes variées sur le cortex supérieur, contenant les deux symbiotes densément associés ;
- des soralies, fissures du thalle permettant l'apparition de sorédies (amas de cellules algales entourées d'hyphes) ;
- des sorédies (sans formation de soralies) ;
- des pycnides (conceptacles à spores).
La reproduction asexuée est assurée par le mycobionte seul qui produit des conidies à l'extrémité des hyphes toujours enfoncés dans le thalle.
La reproduction sexuée, assurée aussi par le mycobionte, forme deux types d'organes spécialisés, les apothécies (forme de cupules à la surface du cortex) ou les périthèces (forme d'outres plus ou moins enfoncées dans le thalle) qui libèrent des spores par rupture du sommet de l'asque selon des mécanismes spécifiques ou par désintégration de la paroi (souvent du type diable en boîte sous l'action de la pression osmotique qui augmente<ref group=alpha>Une partie du glycogène est probablement transformée en glucose, composé qui fait augmenter cette pression osmotique.</ref>, ce qui déchire l'exoascus et fait jaillir l'endoascus comme un ressort)<ref>Modèle:Ouvrage.</ref>. Les apothécies présentent une grande diversité de taille, de couleur et de localisation sur le thalle<ref>Modèle:Ouvrage</ref>.
Ces organes forment un ensemble de caractères très utilisés pour la détermination des espèces.
-
Le thalle du Modèle:Lien est recouvert, à l'exception de la marge, de nombreuses soralies vertes irrégulières.
-
Thalle de Modèle:Lien recouvert d'apothécies enfoncées dans des verrues fructifères sorédiées-pulvérulentes.
{kind=link}
_Flotow_416513.jpg){kind=link}
Nutrition et croissance
.jpg){kind=link}
{kind=link}
{kind=link}
Le mycobionte fournit le support et la protection, les sels minéraux, la réserve d'humidité (une pression osmotique élevée, assurée par l'arabitol et le mannitol, limite la dessiccation du thalle) et facilite probablement l'alimentation du photobionte en Modèle:Formule chimique ; le photobionte fournit les nutriments issus de la photosynthèse chlorophyllienne (amidon chez la plupart des espèces, réserve lipidique chez les Trentepohliales), 20 à 30 % des nutriments étant en moyenne rétrocédés au mycobionte<ref name="Martin">Modèle:Lien web.</ref>.
Les besoins en sels minéraux des lichens sont assez limités, car ce sont de faibles consommateurs qui peuvent provisoirement stopper leur croissance.
Ils se nourrissent à partir de l'atmosphère (minéraux sous forme de solutés dans les eaux de pluie). Les lichens ont aussi la possibilité de dissoudre des éléments minéraux du substrat en excrétant, par l'intermédiaire du champignon, des acides organiques.
Certains sont fixateurs de l'azote atmosphérique (espèces nitrophiles), notamment les cyanolichens.
Ils sont de croissance très lente. En moyenne, la croissance annuelle est de 0,5 à Modèle:Unité pour les lichens crustacés, de 0,5 à Modèle:Unité pour les lichens foliacés et de 1,5 à Modèle:Unité pour les lichens fruticuleux<ref>Modèle:Ouvrage</ref>. Une même espèce peut cependant présenter des taux de croissance très différents en fonction des facteurs environnementaux (climat, nature du substrat, pollution…). Les lichénologues considèrent qu'une croissance de Modèle:Unité est élevée<ref>Modèle:Article.</ref>. Cette caractéristique explique que les lichens résistent peu à la compétition avec des végétaux à développement plus rapide, lesquels inhibent la croissance des espèces pionnières. Elle rend compte aussi de la longévité des lichens qui vivent souvent très longtemps. S'il existe quelques espèces qualifiées d'éphémères (plusieurs du genre Modèle:Lien ne vivent pas plus d'une année), beaucoup sont capables de survivre plus de Modèle:Unité<ref>Modèle:Ouvrage.</ref>, par exemple des alpins<ref>Modèle:Ouvrage.</ref>. L'espèce Rhizocarpon geographicum vit jusqu'à Modèle:Unité dans des zones froides et arides<ref>Modèle:Ouvrage.</ref>. Cette croissance lente et cette longévité sont utilisées par la lichénométrie.
Écologie
Répartition et habitats
{kind=link}
{kind=link}
Cosmopolites et ubiquistes, les lichens recouvrent près de 8 % de la surface terrestre de la planète<ref>Modèle:Article.</ref>. Ils forment les communautés d'espèces autotrophes dominantes dans les écosystèmes polaires et subpolaires<ref>Modèle:Ouvrage.</ref>, où la photosynthèse de certains lichens comme Cladonia alcicornis est toujours effectuée à des températures avoisinant Modèle:Unité<ref>Modèle:Chapitre.</ref>. Plusieurs espèces sont pionnières, capables de coloniser des milieux extrêmes. Elles peuvent s'installer sur des roches qu'elles corrodent en sécrétant des Modèle:Lien (expression souvent inappropriée car nombre de ces molécules sont des phénols qui présentent une fonction acide carboxylique)<ref>Modèle:Article.</ref>. Leur action favorise la succession par des bryophytes, puis par d'autres plantes supérieures<ref>Modèle:Ouvrage.</ref>.
Les lichens ont colonisé pratiquement tous les milieux, Modèle:Citation. Ils sont décrits en fonction de leur substrat<ref>Modèle:Harvsp.</ref> :
- épiphyte (espèce corticole/ramicole sur les arbres/branches, voire sur le bois mort : espèce lignicole) et même follicole (vivant sur les feuilles, particulièrement des plantes sempervirentes) ;
- saxicole : épilithes vivant sur les rochers siliceux ou calcaires (notamment sur les blocs erratiques<ref>Modèle:Article. On y a notamment mis en évidence 16 lichens signalés pour la première fois dans le canton de Vaud en 2020.</ref>), mais qu'on retrouvera sur les vieux murs, les tuiles, ardoises, lauzes ou tôles amiante-ciment, voire sur des supports plastiques ou métalliques (localisation selon la nature et la cohérence de la roche, la pente du rocher, les parois d'orientation, l'exposition aux pluies et écoulements d'eau, l'humidité de la surface déterminée par des suintements Modèle:Incise ou des condensations atmosphériques, la position relative de l'arbre selon qu'il soit isolé ou en massif, en situation ouverte ou fermée…)<ref>Modèle:Article.</ref> ; endolithes vivant dans les rochers ; crypto-endolithes vivant sous les rochers ;
- corticole (espèce qui se développe sur les écorces des troncs, des branches, mais ne les parasite pas). De nombreux facteurs conditionnent l'installation de l'espèce : l'âge de l'arbre et le type d'essence qui influent sur la nature chimique de l'écorce (pH, composition chimique), la texture de l'écorce (crevassée, rugueuse ou lisse) et sa capacité à s'exfolier qui déterminent le pouvoir de rétention de l'eau ;
- terricole et humicole (qui poussent dans pelouses, landes, bois clairs, à l'abri de la compétition des plantes à fleurs<ref>Modèle:Ouvrage.</ref>) ;
- muscicole (sur les mousses) ;
- lichénicole (commensaux ou parasites sur d'autres lichens) ;
- préférence pour les substrats enrichis en dépôts de poussières atmosphériques (espèce coniophile qui trouve dans cette source l'azote ou le phosphate dont elle a plus besoin que d'autres) apportées notamment sur les bords des routes, dans le bas des arbres, sur les toitures (espèce stégophile) ;
- micro-lichens sur les coquilles de patelle ;
- omnicole (espèce non inféodée à un substrat particulier mais qui est capable de se développer sur les substrats naturels et les supports anthropiques les plus divers)Modèle:Sfn.
-
La lécanore des murs est présente sur les routes et les trottoirs. Espèce Modèle:Lien, elle est parfois confondue à un chewing-gum écrasé, résistant comme lui au piétinement<ref>Modèle:Ouvrage.</ref>.
-
Le Modèle:Lien<ref group=alpha>Nom en référence à ses apothécies linéaires, appelées, lirelles (du latin lira, « sillon ») chez certains Ascolichens dits lirellés, apothécie allongée ressemblant à une fente plus ou moins ramifiée à la surface. Le Lichen graphique est doté de lirelles qui dessinent des lignes sinueuses ramifiées évoquant une écriture ancienne.</ref> vit préférentiellement sur les feuillus à écorce lisse, principalement dans la partie inférieure du tronc, en milieu forestier plus ou moins ombragéModèle:Sfn.
{kind=link}
Communautés lichéniques
.jpg){kind=link}
L'étude des peuplements lichéniques commence peu de temps après celle des associations phanérogamiques : les premiers travaux de lichénosociologie datent de 1924, avec le lichénologue polonais Modèle:Lien qui transpose les méthodes d’études des cortèges floristiques aux associations lichéniques<ref>Modèle:Ouvrage.</ref>. Actuellement, la phytosociologie qui étudie les communautés lichéniques, les regroupe en classes, ordres, alliances et associations.
Comme le soulignent Iserentant et de Sloover en 1976<ref>Modèle:Article.</ref>, Modèle:Citation.
Successions de peuplement
Modèle:Section à sourcer Les successions de peuplement typiques de lichens peuvent se résumer schématiquement ainsi : colonisation du substrat (rocher, écorce) par des lichens à thalle crustacé très réduit (endolithique des roches calcaires, épilithique des roches siliceuses, endophléode dans le cas des écorces), puis substitution par des lichens crustacés à thalle plus développé (épilithique ou épiphléode) auxquels succèdent des thalles placodiomorphes (lobés au pourtour) ou squamuleux. Ces derniers sont relayés par de petits lichens foliacés qui, à leur tour sont évincés par de grands foliacés, puis par des fruticuleux et enfin par des bryophytes<ref>Modèle:Article.</ref>.
-
Lepraria incana, lichen sciaphile et Modèle:Lien<ref group=alpha>Lepraria incana se développe dans des endroits ombragés (sciaphile) et abrités des précipitations directes (ombrophobe), sous des rochers acides en surplomb ou à la base des arbres âgés, dans les cavités non directement exposées à l'écoulement des pluies.</ref>.
-
Lecanora carpinea sur rhytidome lisse de troncs, branches et branchettes de feuillus.
-
Xanthoria parietina est un lichen nitrophile<ref group=alpha>La pollution azotée d'origine automobile et agricole modifie la composition spécifique des communautés lichéniques, en favorisant le développement de quelques espèces nitrophiles Modèle:Lien, sans valeur patrimoniale (Xanthoria sp., Physcia sp.). Voir Modèle:Harvsp.</ref> qui a tendance à recouvrir Caloplaca marina et Verrucaria maura, ce qui témoigne d'une croissance plus rapide.
-
Flavoplaca citrina, anciennement Caloplaca citrina, espèce collective très commune sur substrats calcaires naturels ou artificiels.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Résistance aux conditions extrêmes
{kind=link}
Les lichens ont la capacité de résister à de très fortes dessiccations (phénomène de poïkilohydrie). Certains lichens peuvent vivre avec une teneur en eau de 15 %<ref name="Nash_92">Modèle:Harvsp.</ref>. Ils sont aussi capables de se réhydrater (faculté de reviviscence)<ref>Modèle:Harvsp.</ref>, l'absorption hydrique pouvant être telle que les phycolichens renferment jusqu'à 250 à 400 % d'eau, 600 à 2500 % (voire plus) chez les cyanolichens<ref name="Nash_92" />.
La résistance hydrique des lichens provient surtout du mycobionte qui sécrète des polysaccharides autour de l'hyphe, créant ainsi une zone qui piège l'eau sous forme colloïdale. De plus, les lichens accumulent des polyols, qui servent de réserve d'eau. La reprise du métabolisme après une sécheresse est très rapide. Le lichen retrouve ses capacités métaboliques de cinq à trente minutes après une réhydratation<ref>Modèle:Article.</ref>.
Les lichens peuvent également survivre à des variations de température importantes : des tests en laboratoire montrent leur résistance à de hautes températures (Modèle:Unité), à l'azote liquide (Modèle:Unité)<ref>Modèle:Article.</ref>. Ils sont ainsi des constituants majeurs des déserts froids de l'AntarctiqueModèle:Sfn.
En 2005, deux espèces de lichens ont été envoyées dans l'espace et exposées au vide durant deux semaines. Les résultats montrent que, de retour sur Terre et après réhydratation, les lichens survivent à ces conditions extrêmes (dessiccation, températures très basses, rayons UV intenses et rayonnements ionisants) et qu'ils ne présentent quasiment aucune altération de leur structure par rapport à des lichens témoins restés sur Terre<ref>Modèle:Article.</ref>.
Cette résistance à certains stress biotiques et abiotiques (défense contre les agents pathogènes, capacité de supporter de grands écarts de températures ou d'humidité, capacité de bioaccumuler certains métaux lourds et radioéléments dans leur thalle, capacité de dégrader des parties vieillissantes du thalle…), associée à leur autotrophie, fait des lichens des organismes pionniers par excellence<ref>Modèle:Ouvrage.</ref>.
Interactions biologiques
Consommateurs de lichens
{kind=link}
Les chenilles des papillons mycophages suivants se nourrissent de lichens<ref>Modèle:Chapitre</ref>
- la Lithosie aplatie (Eilema complana, Erebidae) ;
- la Lithosie plombée (Eilema lurideola, Erebidae) ;
- le Manteau jaune (Eilema sororcula, Erebidae) ;
- la Ménagère (Dysauxes punctata, Erebidae) ;
- la Rosette (Miltochrista miniata, Erebidae) ;
- la Boarmie des lichens (Cleorodes lichenaria, Geometridae) ;
- la Bryophile vert-mousse (Cryphia algae, Noctuidae).
D'autres animaux, des mammifères tels le yack ou le renne, consomment des lichens<ref>Modèle:Article.</ref>.
Parasites de lichens
Certains champignons ne vivent que sur les lichens en tant que parasites obligatoires. Ceux-ci sont appelés champignons lichénicoles et sont une espèce différente du champignon vivant à l'intérieur du lichen ; ils ne sont donc pas considérés comme faisant partie du lichen<ref name="LFIEB">Modèle:Article.</ref>.
Mimétisme
Intérêt écologique
{kind=link}
Dans l'écosystème, les lichens sont une des composantes parfois importante de la biodiversité<ref>Modèle:Article.</ref>.
Ils sont aussi une source importante de nourriture pour de nombreuses espèces, notamment les rennes. Certaines chenilles de papillons nocturnes, par exemple Noctua promissa, Noctua sponsa, Noctua nupta et Catocala nupta, étaient autrefois dites « lichenées » ou « likenées » parce qu'elles se nourrissent des lichens qui poussent sur les arbres<ref>Modèle:Ouvrage.</ref>.
Ils jouent aussi un rôle important en captant les particules de l'air et des pluies, contribuant à l'épuration permanente des milieux et au recyclage des éléments. Ils se nourrissent de ce que leur apportent l'air et les eaux météoriques et les particules, excréments, pollensModèle:Etc., qu'ils peuvent intercepter et capter. Ils sont capables de faire des réserves et d'accumuler des composés minéraux, bien au-delà des besoins de leur organisme. Cette bioaccumulation est extracellulaire et se fait par le mycobionte. Cela présente des avantages (Modèle:Ex réserve d'éléments comme le phosphore), mais également des inconvénients, comme l'accumulation d'éléments toxiques, voire de radionucléides (par exemple après les essais nucléaires dans l'air ou après la catastrophe de Tchernobyl).
Ils permettent, dans certaines conditions, d'évaluer la chimie et la stabilité des sols, la hauteur moyenne de l'enneigement, l'âge des moraines et le recul des glaciers (datation par lichénométrie), le type de gestion forestière, la durée de la continuité d'un état forestierModèle:Note ; la quantité de polluants dans un milieu donné (les lichens concentrent notamment les métaux lourds<ref group=alpha>Différentes voies sont généralement décrites pour expliquer la prise en charge des contaminants (métaux, radionucléides) et leur bioaccumulation : adsorption sur la surface des parois cellulaires des hyphes fongiques, transport dans les tissus intra-cellullaires, complexation intra-cellulaire à certaines métalloprotéines, complexation extra-cellullaire aux macro-molécules des parois du champignon, complexation avec les acides organiques comme les oxalates ou bien les acides lichéniques. Cf Modèle:Article</ref> — plomb, zinc, cadmium, fluor — et certains radioéléments ou acides dissous dans l'humidité atmosphérique, ce qui peut entraîner leur mort), et surtout le degré de pureté de l'atmosphère (évalué par l'IAP Modèle:Incise prenant en compte le nombre, la polluotolérance et la fréquence des lichens sur un site donné, et l'ILD Modèle:Incise relevant les lichens épiphytes sur les quatre faces des arbres à l'aide d'une grille d'observations)<ref>Modèle:Ouvrage.</ref>.
C'est au biologiste britannique Modèle:Lien que l'on attribue d'avoir observé le premier la diminution des lichens de la région contaminée du Lancashire en 1859<ref>Modèle:Ouvrage.</ref>. Peu de temps après qu'il a mis en évidence le lien entre le déclin des lichens et la qualité de l'air dans la région de Manchester, le botaniste finlandais William Nylander, français d'adoption, décrit Modèle:Nombre et remarque dès 1862 que plusieurs espèces de lichens épiphytes régressent à l'approche des villes, notamment de Paris (1866). Faisant le rapport entre ce déclin et l’usage du charbon, il met en place des bioindicateurs de la qualité de l'air. Depuis, de nombreux lichens sont utilisés en biosurveillance comme indicateurs de pollution<ref>Modèle:Ouvrage.</ref> ou comme informant sur l'origine des certaines pollutions<ref>Modèle:Article</ref>. Les méthodes les plus utilisées pour étudier l'impact des pollutions sur les lichens sont les expériences de fumigation (études de la dégradation de la chlorophylle, de la photosynthèse et de la respiration, bioaccumulation…) et les études de gradient (apparition/disparition de lichens selon un gradient de pollution croissant en fonction de leurs sensibilités respectives à la qualité de l'air)<ref>Modèle:Ouvrage.</ref>.
Une carte de répartition des lichens et des associations lichéniques (technique de phytosociologie) apporte des éléments sur la localisation de zones plus ou moins polluées. Entre 1986 et 1988, deux lichénologues français, Chantal Van Haluwyn et Michel Lerond, mettent au point une méthode phytosociologique qualitative reposant sur l'appauvrissement de groupements lichéniques sous l'action de la pollution. Le travail statistique de leurs relevés les conduit à retenir une trentaine d'espèces de lichens bioindicateurs plus ou moins sensibles aux polluants, répartis en sept zones ou catégories de pollution basées sur une association lichénique<ref>Modèle:Lien web</ref>. La disparition actuelle en ville de certains lichens sensibles notamment au dioxyde de soufre montre que la pollution acide a diminué ; des espèces nitrophiles (lichens orange ou gris sur les arbres) se développent au détriment des espèces neutrophiles ou acidophiles (nitrophobes), montrant l'augmentation de la pollution par les oxydes d'azote. En zone très polluée, on trouve surtout des lichens crustacés, alors qu'en zone moyennement polluée on a surtout des fruticuleux et en zone peu polluée, essentiellement des foliacés et des fruticuleuxModèle:Sfn. Des corrélations ont été observées entre le taux d'azote dans les lichens nitrophiles et l'incidence du trafic routier ou la distance à une usine polluante<ref>Modèle:Article.</ref>.
-
-
Dans les zones de pollution extrême (méthode Van Haluwyn-Lerond), les parties de troncs sont dépourvues de lichens et recouvertes d'une algue microscopique du genre Pleurococcus.
-
Ramalina farinacea, espèce toxicotolérante dont les spécimens de zones polluées sont plus courts et vert foncé.
-
Parmelia sulcata utilisée en biosurveillance pour ses capacités accumulatrices de métaux, de radioéléments et de polluants organiques.
{kind=link}
{kind=link}
{kind=link}
Services écosystémiques
Alimentaires
{kind=link}
{kind=link}
{kind=link}
Certains lichens contiennent des macromolécules de lichénine pouvant être dégradées en glucose au cours de la digestion, ce qui explique que de nombreux lichens ont été utilisés comme aliment de famine<ref>Modèle:Ouvrage.</ref> :
- alimentation animale : les lichens des rennes (Cladonia rangiferina, Cladonia stellaris…) sont la nourriture essentielle du renne et du caribou<ref>Modèle:Article.</ref>. Au Canada, d'immenses troupeaux de caribous lui doivent leur survie, durant les longs mois d'hiver. En toundra ou en forêt tempérée du Nord (où les espèces de lichens sont dix fois plus nombreuses que les plantesModèle:Référence souhaitée), c'est une nourriture appréciée du caribou, de l'élan d'Amérique (orignal) et de certains bétails à l'occasion. Plusieurs variétés de lichens sont comestibles pour eux (notamment Cetraria islandica qui contient 60 % de glucidesModèle:Référence souhaitée), juste un choix de préférence, souvent dû pour certains aux choix de pâturage, aux saisons.
En Suisse, Evernia divaricata a été longtemps utilisé pour nourrir les chèvres en mauvaise saison ; - alimentation humaine : certains peuples nordiques consomment la mousse d'Islande (Cetraria islandica) en farine pour en faire des pains ou gâteaux. Divers Umbilicaria sont également consommés au Canada (les « tripes de roche »); Lasallia pustulata est également appelée « tripe de roche », ce lichen saxicole calcifuge ayant longtemps été utilisé comme aliment de famine<ref>Modèle:Ouvrage.</ref>. Au Japon, Umbilicaria esculenta est consommé en soupe, en tenpura et en salade sous le nom de Iwatake イワタケ (岩竹, « bambou des roches »). Plusieurs espèces du genre Modèle:Lien ou Lecanora (notamment Modèle:Lien) sont parfois identifiées à la manne des Hébreux<ref>Modèle:Article.</ref>. Parmotrema perlatum est utilisé comme épice dans la cuisine de certaines régions de l'Inde, en particulier la cuisine chettinad du Tamil Nadu. La plupart des espèces de lichens sont considérées comme « comestibles » à condition de les faire macérer et bouillir pour en retirer les composés acides et amers. Quelques espèces (Cetraria pinastri, Letharia vulpina, Xanthoparmelia chlorochroa dont le simple contact peut entraîner des réactions allergiques) sont très toxiques car elles contiennent de l'acide vulpinique, toxine qui agit en stoppant l'activité respiratoire. L'ébullition ne permet d'extraire qu'une partie de cette toxineModèle:Sfn ;
- enfin, quelques espèces sont utilisées comme émulsifiant et épaississant dans l'industrie agroalimentaire.
Médicinaux et médicamenteux
{kind=link}
_i_Skymlag%C3%A5rdens_mark,_S%C3%B6rby_sn_2619.jpg){kind=link}
Modèle:Unité de lichens trouvent des applications en médecine traditionnelle<ref>Modèle:Chapitre</ref>.
{kind=link}
Si cette pharmacopée est aujourd'hui tombée dans l'oubli et certaines croyances des anciens font désormais sourire, la recherche médicale moderne (notamment en industrie pharmaceutique et en microbiologie) met en évidence que de nombreux lichens représentent une source prometteuse de produits naturels originaux pour la recherche de nouvelles molécules bioactives ayant un intérêt pharmaceutique (activité antioxidante, antimicrobienne, antifongique, antivirale, anticancéreuse, anti-inflammatoire, insecticide, photoprotectrice ou photostimulatrice<ref>Modèle:Article.</ref>,<ref>Modèle:Article.</ref>,<ref>Modèle:Article.</ref>. Cette source comprend des composés endolichéniques d'origine fongique mais aussi d'origine bactérienne (les lichens constituant une niche écologique pour de nombreux micro-organismes)<ref>Modèle:Article.</ref>. Cependant, la plupart des recherches étudiant les effets thérapeutique de ces composés, ne parviennent pas à cibler les molécules pouvant être responsables des propriétés bénéfiques pour la santé, ce qui explique qu'elles n'atteignent pas les étapes finales de la découverte de médicaments à des fins thérapeutiques<ref>Modèle:Article.</ref>. Ces composés correspondant à certains des 1 100 métabolites secondaires lichéniques<ref>{{#invoke:Langue|indicationDeLangue}} John A Elix, Catalogue de substances lichéniques, 3e édition, Australian National Botanic Gardens, 2014</ref>, molécules à 90 % originales produites par trois voies métaboliques qui correspondent à l'équipement enzymatique du mycobionte (voie des polyacétates appelée aussi voie acétate-polymalonate, voie du mévalonate, voie du shikimate)<ref>Modèle:Article.</ref>.
Exemples d'applications :
- Se basant sur la théorie des signatures, la médecine traditionnelle utilisait des espèces comme le lichen d'Islande (Cetraria islandica), la pulmonaire (Lobaria pulmonaria dont le thalle a la forme de poumons), le lichen pyxidé (Cladonia pyxidata) autrefois réputés pour la toux, et le lichen du chien (Peltigera canina). L'usnée du crâne humain (Usnea plicata) récoltée sur le crâne des pendus, se payait à prix d'or pour guérir, croyait-on, l'épilepsie ;
- des extraits d'Umbilicaria esculenta se sont révélés fortement inhibiteurs de la β-glucosidase, les enzymes qui hydrolysent les disaccharides chez les moisissures et les cellules de mammifères, ainsi que la glucoamylase et de la laminarinase. Elle a des propriétés thrombolytiquesModèle:Référence nécessaire..
Les propriétés antimicrobiennes expliquent que des lichens comme Pseudevernia furfuracea ont été utilisés dans les processus de momification en Égypte antique<ref>Modèle:Article.</ref>
Industrie teinturière
{kind=link}
Les lichens sont utilisés en teinture végétale<ref>Modèle:Ouvrage.</ref>. On tire des acides lichéniques des colorants de nuances assez riches. De belles teintes de bleu, pourpre (rouge violacé) et violet sont données par la parelle d'Auvergne, Ochrolechia parella, l'orseille des Canaries (Roccella tinctoria qui, extrait en poudre, produit le papier de tournesol ou le colorant alimentaire orcéine). D'autres espèces sont utilisées traditionnellement, notamment en Scandinavie :Candelaria, Rocella phycopsis, Rocella fuciformis, Ochrolechia tartarea, Pertusaria dealbescens, Parmelia glabratula subsp. fuliginosa et Lasallia pustulata. Les Xanthoria pilés et mélangés à de l'urine servaient à teindre la laine en rose, notamment du kilt écossais<ref>Modèle:Ouvrage.</ref>. La teinture traditionnelle de ces kilts obtient encore aujourd'hui différentes teintes selon l'espèce de lichen et le temps de décoction dans l'eau bouillante<ref>Modèle:Ouvrage.</ref>.
Parfumerie
{kind=link}
.jpg){kind=link}
Certains lichens sont utilisés pour produire des huiles essentielles de parfum. L'industrie parfumière tire notamment parti de deux lichens fruticuleux récoltés sur les arbres : la mousse des arbres (Pseudevernia furfuracea, corticole des conifères, que l'on trouve souvent au sol, détachée par le vent) qui apporte des notes boisées variées en fonction de son arbre support, et la mousse de Chêne (Evernia prunastri) qui apporte des notes marines utilisées dans les parfums féminins. On extrait de ces lichens un produit fondamental en parfumerie, l'absolue mousse considérée comme une note de fond, capable de produire des notes boisées avec des nuances de champignons et d'algues<ref>Modèle:Harvsp.</ref>. Près de Modèle:Unité de ces deux espèces étaient récoltées annuellement dans la région de Grasse, des Balkans et au Maroc<ref>Modèle:Article.</ref>. Cependant, cette récolte décline en raison de la présence dans ces extraits de composés allergènes, notamment l'atranorine<ref>Modèle:Ouvrage.</ref>. Ces composés bioactifs sont ainsi de plus en plus remplacés par des produits synthétiques<ref>Modèle:Article.</ref>.
Autres
Durant la préhistoire, les mèches utilisées pour l'allumage des lampes à graisse sont des champignons (amadouvier), des algues, des mousses séchées, mais aussi des lichens fruticuleux séchés prélevés le plus souvent sur des branches d'arbres ou d'arbustes (tel qu'Evernia prunastri) ou au sol<ref>Modèle:Article.</ref>.
Toxicité
On a prétendu jadis qu'aucun lichen n'était vraiment vénéneux<ref>Modèle:Ouvrage.</ref>, à l'exception de quelques troubles intestinaux provoqués par les espèces très amères. Par la suite, on a rapporté l'utilisation de Letharia vulpina (corticole des mélèzes ou pins alpins, sa couleur vert-jaunâtre est due à l'acide vulpinique, toxique utilisé comme appât pour tuer les loups et renards) et Cetraria pinastri en Scandinavie pour empoisonner les loups. Le principe toxique agirait par inhibition de la respiration, entraînant la mort<ref>Henry des Abbayes (1931) - Traité de lichénologie, p. 185</ref>.
L'orseille extraite (voir au chapitre précédent) a été interdite comme colorant alimentaire à cause de sa toxicité.
Depuis, comme pour les champignons, la liste des lichens toxiques ne cesse de s'allonger, parmi lesquels :
- Letharia vulpina, Cetraria pinastri, Xanthoparmelia chlorochroa.
Histoire évolutive
{kind=link}
Les partenaires de la symbiose ont d'abord existé sous forme d'espèces libres dans le milieu marin : bactéries, algues cellulaires à la superficie des eaux et microchampignons aquatiques. La conquête des terres émergées par des bactéries au cours de l'Archéen, puis par des algues et des champignons microscopiques au Protérozoïque, se traduit initialement par la colonisation des pentes marécageuses puis par la terre ferme. Au cours de l'évolution, des espèces fongiques exploitent des ressources fournies par un autre individu non apparenté (algues ou cyanobactéries), au détriment de celui-ci, développant un parasitisme nécrotrophe : des champignons surexploitent les ressources de leur partenaire au point de le faire mourir, se nourrissant de ses restes avant de capturer de nouveaux individus. Ces espèces ont pu aussi établir un autre mode de relation trophique, le parasitisme biotrophe, interaction biologique stable lorsque l'individu parasité élabore des mécanismes de défense suffisamment puissants pour éviter sa mort mais pas assez pour éliminer le parasite. Cette symbiose parasitaire a pu évoluer en symbiose mutualiste qui, de facultative devient obligatoire à mesure que le degré de dépendance entre les deux partenaires augmente<ref>Modèle:Ouvrage.</ref>. En effet, une vraie conversion parasitisme-mutualisme est possible si des mutations favorables à la symbiose se produisent dans le génome de l'espèce parasite ou parasitée. L'exemple typique est la mutation délétère de gènes fabriquant des protéines produites par le partenaire, ce qui permet une économie métabolique. L'inactivation de ces gènes devenus inutiles, entraîne leur disparition du génome à l'occasion de délétions ou leur transformation en pseudogènes, et engage les deux partenaires sur une voie évolutive sans retour, la dépendance<ref>Modèle:Ouvrage.</ref>.
L'enregistrement fossile de lichens est rare<ref>Modèle:Article.</ref>. Les premiers fossiles connus sans ambiguïté ne remontent pas avant la colonisation des terres par les plantes, si bien qu'il n'y a aucune preuve de l'émergence des lichens avant l'évolution des plantes terrestres<ref>Modèle:Article.</ref>.
Les lichens forment un groupe polyphylétique<ref>Arbre phylogénétique des champignons. Le mode de vie symbiotique de type lichen est indiqué en rouge dans la seconde colonne. Cf Modèle:Article.</ref>. En d'autres termes, les processus de lichénisation sont apparus plusieurs fois indépendamment dans l'histoire évolutive des champignons et de leurs symbiotes. Malgré leurs origines multiples, ils présentent un mode de vie et des adaptations communes (taille réduite, production de métabolites secondaires<ref group=alpha>Près de Modèle:Unité (terpénoïdes, dérivés d'orcinol, chromones, xanthones, depsides, depsidones) ont été recensés, la plupart étant exclusivement synthétisés par les lichens. Cf Modèle:Chapitre</ref> aux rôles divers Modèle:Incise, capacité de déshydratation et de vie ralentie en cas de manque d’eau puis de reviviscence reviviscence par réhydratation), illustrant le phénomène de convergence évolutive<ref>Modèle:Ouvrage.</ref>.
{kind=link}
Ainsi, selon Marc-André Selosse, Modèle:Citation.
Systématique
On donne des noms scientifiques séparés à chaque champignon et à chaque algue qui constituent un lichen, mais pas de nom unique à la combinaison particulière de ces espèces, en tout cas en 2011. Quand on dit qu'un lichen appartient à une espèce, c'est en fait l'espèce du principal champignon qui le constitue, même si celui-ci forme des lichens d'apparences différentes avec plusieurs espèces d'algues séparément<ref name="Kirk378_381">Modèle:Ouvrage.</ref>,<ref name=FSSD>Modèle:Lien web.</ref>.
La classification d'Alexander Zahlbruckner (1860-1938) (1907, 1926), malgré son ancienneté, garde une valeur pratique face aux classifications récentes souvent incomplètes. Elle décompose la classe des lichens selon le schéma suivant :
- Sous-classe des Ascolichenes : spores produites dans des asques.
- Série des Pyrenocarpeae : ascocarpes ne s'ouvrant que par un pore, thalles en général crustacés (environ dix-sept familles).
- Série des Gymnocarpeae : ascocarpes plus ou moins largement ouverts, thalles de tous les types.
- Sous-série des Graphidineae : asques et paraphyses se détruisant et formant avec les spores, dans l'ascocarpe, un amas pulvérulent. Le thalle est en majorité crustacé, ou fruticuleux. Il existe trois familles.
- Sous-série des Cyclocarpineae : ascocarpes de forme arrondie. C'est le groupe le plus nombreux, où se trouvent tous les types de thalles. Il existe vingt-neuf familles.
- Sous-classe des Basidiolichenes (ou Hymenolichenes) : spores produites sur des basides (Omphalina, Modèle:Lien).
Une autre classification est basée sur le composant fongique qui joue le rôle principal dans la détermination de la forme générale du lichen : Modèle:Palette Taxinomie des familles de lichen Les lichens comme les autres espèces sont identifiés au moyen de clés de détermination (Exemple<ref>Modèle:Lien web.</ref>), mais des analyses chimiques ou génétiques, ou l'utilisation d'indicateurs colorés peuvent être nécessaires pour identifier avec certitude certaines espèces ou leur composantes fongiques et algales.
Dans la culture
Dans le calendrier républicain, le Lichen est le Modèle:17e du mois de pluviôse<ref>Modèle:Ouvrage.</ref>.
{kind=link}
En raison de leur taille réduite, de leur discrétion et de leur abondance, les lichens sont rarement représentés dans les arts<ref name="Pacini">Modèle:Chapitre.</ref>.
Les lichens inspirent de nombreux artistes romantiques ou naturalistes, écrivains (Marcel Proust, Rousseau, George Sand), poètes (Henry David Thoreau, Francis Ponge), peintres (Christian Dotremont, Jasper Johns), plasticiens (Robert Rauschenberg, John Cage)<ref>Modèle:Ouvrage.</ref>…
Notes et références
Notes
Références
Voir aussi
Bibliographie
- Modèle:Article.
- Modèle:Ouvrage. Modèle:Plume
- Modèle:Article.
- Modèle:Ouvrage. Modèle:Plume
- Modèle:Ouvrage. Modèle:Plume
- Modèle:Article.
- Modèle:Ouvrage.
- Modèle:Ouvrage.
- Modèle:Article